|
Student Opportunities in the Locomotion Lab
Many of my projects involve mentoring undergraduate students with an interest in research. Students are involved with all stages of a project, from project development and construction of equipment to data collection and analysis to manuscript development and publication. Many students who work in my lab have the opportunity to present their work at conferences and/or co-author peer-reviewed publications. |

|
Current Projects
Squirrel Studies
Urbanization and anthropogenic fragmentation of natural habitats can introduce novel stressors to animals that can significantly influence population size and local biodiversity. One contributing factor to increases in population density in urban environments may be access to supplemental food sources in the form of human food waste, bird feeders, and pet food. Access to supplemental food sources may also promote greater body mass in urban individuals compared to rural counterparts. Such body mass increases may be advantageous for overwintering survival and provisioning offspring, however increases in mass may also have detrimental locomotor consequences. For this work, we are assessing body mass and field locomotor (e.g. climbing and jumping) performance in fox squirrel populations on the campus of Saint Mary's College and in the nearby nature area.
Animal body mass is also know to fluctuate regularly with reproductive cycles. Such annual body mass changes may be particularly drastic in gravid females. Increases in mass are known to affect an organism's center of mass, which can have a substantial effect on individual balance and stability, especially during locomotion. However, tails have been shown to function as stabilizing structures during arboreal locomotion, with longer tails acting as more effective stabilizers compared to shorter tails. Because squirrels inhabit arboreal environments characterized by unstable and discontinuous substrates, annual increases in body mass related to female reproductive state may present special locomotor challenges in this environment. We suspect that female squirrels may possess longer tails compared to males of the same species to counteract impacts of gravity on locomotor stability. For this work, we are comparing male and female tail length in squirrels to determine whether there are sex-specific differences in tail morphology.
Urbanization and anthropogenic fragmentation of natural habitats can introduce novel stressors to animals that can significantly influence population size and local biodiversity. One contributing factor to increases in population density in urban environments may be access to supplemental food sources in the form of human food waste, bird feeders, and pet food. Access to supplemental food sources may also promote greater body mass in urban individuals compared to rural counterparts. Such body mass increases may be advantageous for overwintering survival and provisioning offspring, however increases in mass may also have detrimental locomotor consequences. For this work, we are assessing body mass and field locomotor (e.g. climbing and jumping) performance in fox squirrel populations on the campus of Saint Mary's College and in the nearby nature area.
Animal body mass is also know to fluctuate regularly with reproductive cycles. Such annual body mass changes may be particularly drastic in gravid females. Increases in mass are known to affect an organism's center of mass, which can have a substantial effect on individual balance and stability, especially during locomotion. However, tails have been shown to function as stabilizing structures during arboreal locomotion, with longer tails acting as more effective stabilizers compared to shorter tails. Because squirrels inhabit arboreal environments characterized by unstable and discontinuous substrates, annual increases in body mass related to female reproductive state may present special locomotor challenges in this environment. We suspect that female squirrels may possess longer tails compared to males of the same species to counteract impacts of gravity on locomotor stability. For this work, we are comparing male and female tail length in squirrels to determine whether there are sex-specific differences in tail morphology.






Human Kinematics Studies
Anterior Cruciate Ligament (ACL) is a predominant injury among adolescents and young adults. Such injuries typically require a lengthy recovery period, including multiple physical therapy appointments per week for several month, which often cause students to miss classes and is accompanied by high financial cost. Furthermore, ACL injury predisposes individuals to re-injury and development of secondary disorders like osteoarthritis. Similarly, repetitive stress injuries in other joints can have similar consequences for dancers and musicians. Taken together, the academic, athletic, and financial costs of an joint injury have a substantial negative impact on both short-and long-term quality of life for the injured individual.
The objective of our research is to assess the prevalence of joint injury in college-age students at Saint Mary's College (and neighboring communities) and to identify differences in kinematic variables associated with athletic activity in individuals who have and have not sustained an ACL or other serious knee injury. We are also planning a second project that examines arm joint kinematics and history of repetitive stress injury in string musicians. These data will be employed in the development of an affordable image-capture and software package capable of tracking anatomical positions in 3-dimensional space and identifying neuromuscular and biomechanical deficiencies that may contribute to joint injuries in athletes and musicians.
Anterior Cruciate Ligament (ACL) is a predominant injury among adolescents and young adults. Such injuries typically require a lengthy recovery period, including multiple physical therapy appointments per week for several month, which often cause students to miss classes and is accompanied by high financial cost. Furthermore, ACL injury predisposes individuals to re-injury and development of secondary disorders like osteoarthritis. Similarly, repetitive stress injuries in other joints can have similar consequences for dancers and musicians. Taken together, the academic, athletic, and financial costs of an joint injury have a substantial negative impact on both short-and long-term quality of life for the injured individual.
The objective of our research is to assess the prevalence of joint injury in college-age students at Saint Mary's College (and neighboring communities) and to identify differences in kinematic variables associated with athletic activity in individuals who have and have not sustained an ACL or other serious knee injury. We are also planning a second project that examines arm joint kinematics and history of repetitive stress injury in string musicians. These data will be employed in the development of an affordable image-capture and software package capable of tracking anatomical positions in 3-dimensional space and identifying neuromuscular and biomechanical deficiencies that may contribute to joint injuries in athletes and musicians.

Turtle Studies
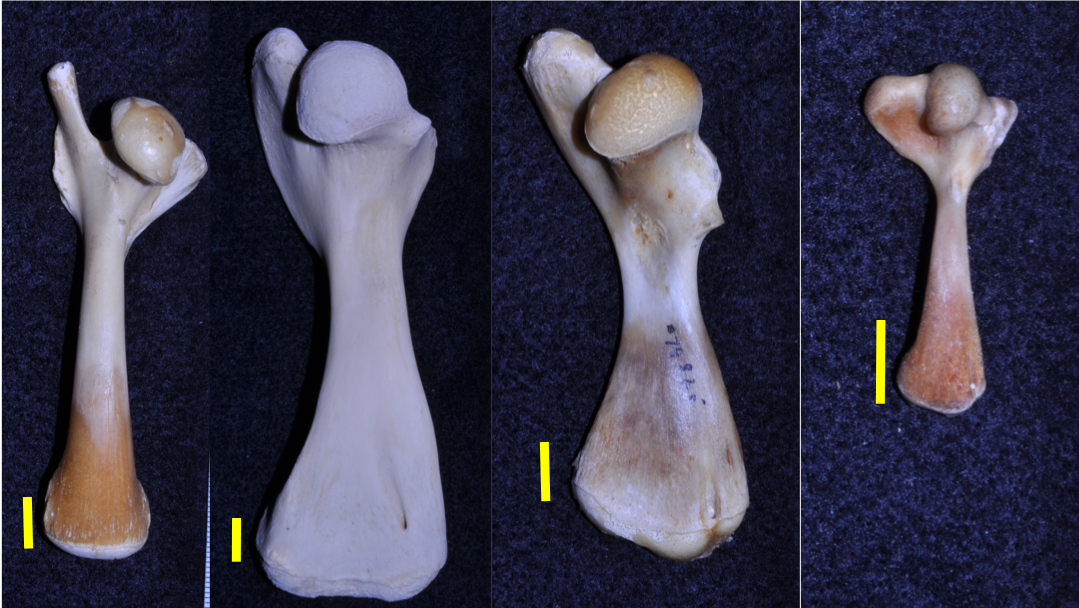
Emydids - Turtle species within the emydid clade span a range of functional and life history strategies. Most members of this clade are semi-aquatic rowers, however four emydid species, all in the genus Terrapene, have evolved into terrestrial specialists from semi-aquatic ancestors. Inclusion of these terrestrial specialists in previous emydid scaling analyses may have skewed results disproportionately toward a terrestrially specialized scaling pattern for this group. Additional analyses of proximal limb bone scaling patterns that separate terrestrail from semi-aquatic emydid taxa are necessary to resolve this possibility.
Pleurodires - Prior limb bone scaling analyses have limited comparisons to cryptodiran taxa. Pleurodiran taxa exhibit aquatic specializations and locomotor strategies similar to some cryptodiran freshwater specialists (e.g. softshell turtles), however patterns of limb bone scaling have been largely overlooked in pleurodiran taxa.
Emydids - Turtle species within the emydid clade span a range of functional and life history strategies. Most members of this clade are semi-aquatic rowers, however four emydid species, all in the genus Terrapene, have evolved into terrestrial specialists from semi-aquatic ancestors. Inclusion of these terrestrial specialists in previous emydid scaling analyses may have skewed results disproportionately toward a terrestrially specialized scaling pattern for this group. Additional analyses of proximal limb bone scaling patterns that separate terrestrail from semi-aquatic emydid taxa are necessary to resolve this possibility.
Pleurodires - Prior limb bone scaling analyses have limited comparisons to cryptodiran taxa. Pleurodiran taxa exhibit aquatic specializations and locomotor strategies similar to some cryptodiran freshwater specialists (e.g. softshell turtles), however patterns of limb bone scaling have been largely overlooked in pleurodiran taxa.

|

|
Salamander Studies
Laterally compressed tails are typically associated with aquatic tetrapods because increased surface area help to generate propulsive thrust in aquatic environments. In contrast, tails that are round in cross-section are more often observed in terrestrial taxa. Preliminary analyses of tail shape and swimming performance in blue-spotted salamanders (Ambystoma laterale) and redback salamanders (Plethodon cinereus) indicate that factors such as tail beat amplitude, swimming gait, and tail beat frequency may have a greater influence on swimming velocity than tail morphology.
Laterally compressed tails are typically associated with aquatic tetrapods because increased surface area help to generate propulsive thrust in aquatic environments. In contrast, tails that are round in cross-section are more often observed in terrestrial taxa. Preliminary analyses of tail shape and swimming performance in blue-spotted salamanders (Ambystoma laterale) and redback salamanders (Plethodon cinereus) indicate that factors such as tail beat amplitude, swimming gait, and tail beat frequency may have a greater influence on swimming velocity than tail morphology.

Previous WorkTetrapod Functional Morphology and Secondary Aquatic Invasions
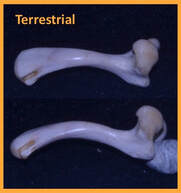
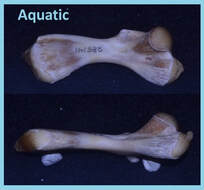
The invasion of land by early tetrapods is arguably one of the most profound events in vertebrate history. However, equally profound but less often studied is the return of terrestrial vertebrates to highly or exclusively aquatic lifestyles. This secondary shift back to the water is often accompanied by characteristic morphological changes, including the flattening of the shaft of the long bones in the limb. Such morphological changes have largely been attributed to intuitive changes in loading regime between land and water, with reduced body support demands in aquatic habitats. These projects investigated potential mechanisms for how flattening of the limb long bones may come about. Femoral Loading: Strain data from the femur of semi-aquatic Trachemys scripta (slider turtle; Family: Emydidae) confirm the expected reduction in load magnitudes during swimming compared to terrestrial walking. In addition, torsional (twisting) loads on the femur were also significantly reduced during swimming compared to walking. In an environment in which torsional loads are not prevalent (e.g. aquatic), animals may be released from the need to maintain a round cross-sectional shape, thus permitting the shift to a more hydrodynamically advantageous flattened shape of the long bones. Humeral Loading: Data from the humerus of the semi-aquatic Pseudemys concinna (river cooter turtle; Family: Emydidae) show similar patterns to those observed in the slider femur. These data indicate that changes in terrestrial and aquatic loading regime are associated more with a reduction in overall strain magnitude rather than changes in load orientation (change in torsional loads). Thus, despite similar reductions in shear strain between terrestrial and aquatic locomotion, these reductions are produced by different mechanisms. Swimming Kinematics: Specialized performance in a particular habitat often comes at the cost of high performance in contrasting habitats. However, retention of ancestral performance ability in specialist species has been largely unexplored. Despite aquatic ancestry, some turtle lineages have transitioned to exclusively terrestrial habits. Comparison of swimming kinematics of the forelimb and hindlimb of terrestrial tortoises (Testudo horsfieldii) and box turtles (Terrepene carolina) to two semi-aquatic emydid turtles (Trachemys scripta and Chrysemys picta) indicates functional convergence of the forelimb in box turtles and tortoises, but the box turtle hindlimb retains ancestral swimming motions. Phylogenetic Limb-Bone Scaling: Although strain data from turtle humeri and femora indicate a potential mechanism for how flattened limb bone shapes may be achieved, such a mechanism is difficult to assess without considering bone morphology across the functional spectrum exhibited in turtles (including terrestrial walkers, aquatic rowers, and marine flappers/underwater flight). Data were collected from skeletal specimens housed at the American Museum of Natural History, the Carnegie Museum of Natural History, the Florida Museum of Natural History, the Smithsonian National Museum of Natural History, and the Chelonian Research Institute. Limb bone size and shape patterns reflect functional and life history differences among the taxa examined, indicating that limb bone shape is likely governed by a combination of mechanical, ecological, and evolutionary factors. |




|


|
Physiological and Morphological Variation in a Stream Salamander
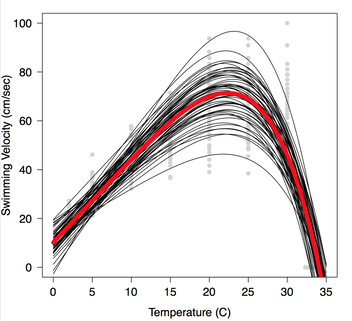
Habitats vary in their biotic and abiotic components across space and time. Such variability can introduce challenges for organisms unless they have the ability to alter physiology and morphology in a way what maximizes fitness. Thermal Acclimation: Theory predicts that highly variable thermal environments should lead to reduced thermal sensitivity in ectotherms, resulting in greater acclimation capacity. Salamanders of the species Desmognathus brimleyorum (Ouachita dusky salamander; Family: Plethodontidae) were exposed to constant and variable thermal conditions in the lab. Following the acclimation period, thermal sensitivity of metabolic rate and swimming performance was assessed across a range of temperatures. Swimming performance did not show a significant difference between treatments (see figure at left showing individual performance curves in black and mean performance in red), however a limited degree of acclimation of metabolic rate was observed. Morphological Variation: Individual salamanders responded to treatments differently depending on which source population they originated from. Furthermore, tail morphology appeared to differ between populations, though sample sizes were too small to statistically confirm this difference. Additional work with this system is necessary to further evaluate the extent of morphological diversity between populations and to determine what environmental elements may be driving these differences in tail shape. |

|
